-
【听课笔记】复旦大学遗传学_05染色体畸变
课程地址:复旦大学遗传学
五、染色体畸变
5.1 染色体分析方法
5.1.1 核型
- 细胞中染色体的总体构成称为核型 (
karyotype)。 - 对标本的染色体构成进行检查称为核型分析 (
karyotyping)。核型分析是细胞遗传学的主要任务。 - 核型分析的一般步骤
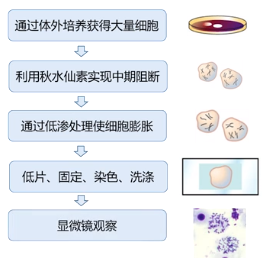
(1) 通过体外培养获得大量细胞:从体内分离细胞,在体外扩大培养;
(2) 利用秋水仙素实现中期阻断:秋水仙素可以和微管结合,破坏纺锤体的作用,使大量细胞停留在分裂中期。
(3) 通过低渗处理使细胞膨胀;
(4) 滴片、固定、染色、洗涤。低渗处理的膨胀细胞悬液滴在载玻片上,细胞就会破碎,释放其中的染色体组,然后固定、染色、洗涤、封片观察。
(5) 显微镜观察。在光学显微镜下可以看到,粗短的染色体相对集中地分布在一个圆形区域,即细胞破碎的地方,同时不同染色体之间能够相互清楚的区分开来。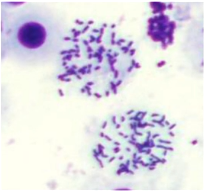
染色体的形态与类型
-
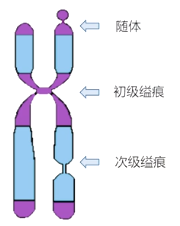
着丝粒的位置:
■ 中央着丝粒染色体:着丝粒在染色体 1/2 ~ 5/8 处的染色体(几乎在中部)
■ 亚中央着丝粒染色体:着丝粒在染色体 5/8 ~ 7/8 处的染色体
■ 近端着丝粒染色体:着丝粒在染色体 7/8 ~ 近末端的染色体

-
次缢痕:
■ 着丝粒所在的地方往往表现为一个缢痕,所以着丝粒又称为初级缢痕。
■ 有的染色体在染色体壁上还有次级缢痕,其位置是固定的,也属于染色体的形态。
■ 在一些染色体末端还连接着一个叫随体的远端染色体小段,也属于染色体的形态。
-
当我们了解了一个细胞内全部染色体的数目、大小和形态时,就可以将全部染色体按照特定顺序排列并合理分组,如此编排的染色体图像,就是核型的常见表示方法。

5.1.2 染色体显带
-
染色体显带技术: 借助特殊的染色体处理方法,然后利用染料将染色体沿长轴染成宽窄及明暗不同的条带,以供染色体的辨别和变异染色体的鉴定的技术方法。
-
常见的显带技术有:G 显带、Q 显带、C 显带和 R 显带。
- G 显带 (
G banding):将染色体经热、碱或蛋白酶等预处理后进行 Giemsa 染色后获得的带型。观察简便,且标本易长期保存。使用最广泛。

- Q 显带 (
Q banding):荧光染料处理染色体后在紫外光激发下形成的特定带型。显带效果好,但观察复杂且不易保存。 - R 显带 (
R banding):用磷酸盐溶液预处理后的染色体标本进行 Giemsa 染色获得的带型。其带型跟 G 带正好相反,可分析染色体 G 带浅带部位的结构改变。 - C 显带 (
C banding):用 NaOH 预处理染色体后的 Giemsa 染色后获得的带型。可以特异性显示着丝粒及染色体内部或端部的异染色质区染色结构。
- G 显带 (
-
果蝇唾腺染色体:
■ 果蝇及其他双翅目昆虫的幼虫的唾腺细胞染色体属于特殊的多线染色体,是染色体分析的好材料。
■ 多线染色体:细胞停留在分裂间期或前期,染色体连续复制,但姐妹染色体单体并不分开,而是纵向聚集在一起,且这些细胞的细胞核和细胞都不分裂。
■ 所以,正常的果蝇染色体是四对染色体,但在唾腺中,染色体非常巨大,且全部的着丝粒及其邻近部分相互聚合形成了染色中心。
■ 染色中心 (chromocenter):四对染色体的着丝粒及其邻近部分相互聚合。
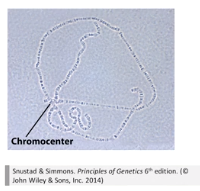
■ 因为染色体螺旋化程度不同,唾腺染色体上还有深浅不一的横纹,它们的相对大小和位置恒定,染色后清晰可见,所以便于对染色体结构变化进行遗传分析。
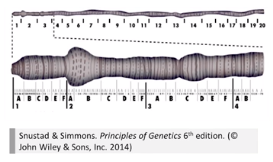
5.1.3 人细胞的核型分析
- 人细胞的核型
1960 年,首届国际细胞遗传学会议,确定正常人的核型基本特征,即丹佛体制 ( Denver system )。丹佛系统根据人类长度及着丝粒的位置将染色体分为 7 组。
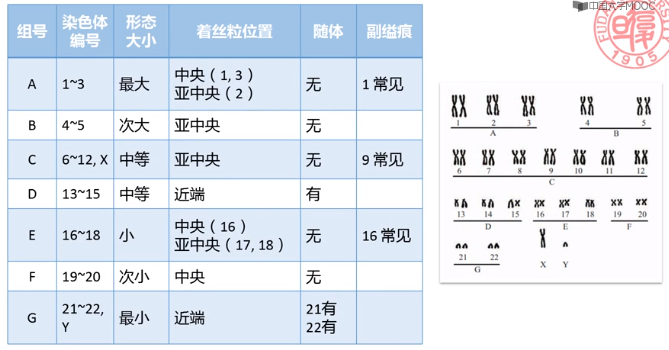
- 人类染色体的描述方法
■ 人类细胞遗传学命名的国际体制 (简称 ISCN ),每条染色体根据 ISCN 规定的界标划分成若干个区、带、亚带和次带。
■ 界标landmark:对于识别染色体有重要帮助的染色体特征,如着丝粒、末端和恒定的带等。
■ 长臂q;短臂p
■ 区region代表的是相邻染色体界标之间的区域
■ 带band是显色技术下可见的条带
■ 亚带和次亚带 是利用高分辨显带技术显示的更加精细的条带
■ 区和带的编号都是从着丝粒开始,两端逐渐变大
■ 描述时需要依次写出染色体号、长/短臂、区号、带号、小数点、亚带号和次亚带号。如下图分别表示的是人类 5 号染色体上短臂 1 区 5 带 3 亚带 2 次亚带、5 号染色体上长臂 1 区 2 带 1 亚带。

5.2 染色体重复与缺失
5.2.1 染色体结构变异
- 染色体畸变 (
chromosomal aberrations) 包括染色体结构和数目的改变。 - 染色体结构变异指的是染色体在结构上的大片段改变,起源于染色体或染色单体的断裂和重接。染色体结构变异包括缺失、重复、倒位、易位、环状染色体、插入、双着丝粒染色体和等臂染色体等多种类型。利用核型分析和显带技术可以识别不同的染色体结构变异。
- 染色体结构变异可以带来不同的细胞学和遗传学效应。
5.2.2 染色体缺失
-
染色体缺失 (
deletion) 是指染色体的片段出现丢失。染色体缺失通常会导致个体的生活力下降。缺失纯合体的生活力比缺失杂合体的生活力更低,因为纯合体丢失了 2 个拷贝的染色体片段。 -
染色体缺失可以分为中间缺失和末端缺失,两者产生的机制略有不同。
-
中间缺失 (
interstitial deletion) 起源于同一条染色体臂上发生了两次断裂, 断点之间无着丝粒片段丢失,其余片段重接。
中间缺失案例:46, XY, del(3)(q21q31)
46 条染色体,性染色体组成为 XY,3 号染色体的 q21 至 q31 区域发生缺失。

-
末端缺失 (terminal deletion) 起源于一条染色体发生一次断裂后未发生重接, 丢失无着丝粒的染色体片段。
末端缺失案例:46, XX, del(4)(q27)
46 条染色体,性染色体组成为 XX,4 号染色体的 q27 至末端区域发生缺失。

-
缺失的细胞学效应
■ 缺失杂合体联会时,缺失的染色体区段和同源染色体的相应区段不能配对,被“拱”起来,出现弧状结构。
■ 缺失造成的弧状结构的内部是正常的染色体部分。
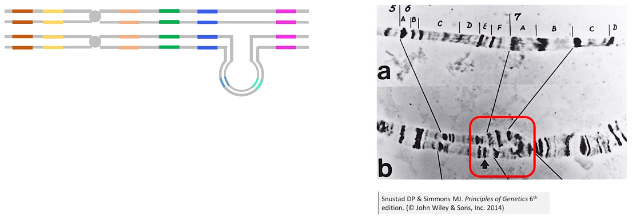
-
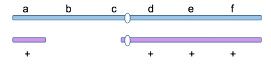
缺失的遗传学效应——拟显性
拟显性(pseudodominance):杂合子中,由于显性基因的缺失,使原来不应表现出来的隐性非致死等位基因的效应显现出来。
-
缺失的遗传学应用——基因定位
-
通过分析不同突变品系的拟显性的表型特征以及发生缺失的染色体区段,推导出表型决定基因所在的染色体位置。
-
案例:想定位一个突变基因——小眼不齐基因 fa,已知它在 X 染色体上,属于隐性突变,同时实验室有 13 种 X 染色体上不同程度的缺失的果蝇品系,红色方框表示各个品系缺失的染色体区域。
■ 将 fa 基因的纯合突变体与各个缺失品系杂交,得到携带 fa 缺失基因的缺失杂合子(雌蝇),如果这些杂合子 fa 基因所对应的位置上存在缺失,这些杂合雌蝇就会表现出拟显性现象(小眼不齐),这个杂合子中的染色体缺失区域一定覆盖了 fa 基因的所在位置,那么只要求出所有出现拟显性现象的果蝇缺失品系缺失区域的最大重叠区间即可,如下图,将 fa 基因定位在了他们共同缺失的 3C7 这个横纹中。

-
案例:猫叫综合征
cri-du-chat syndrome
■ 5 号染色体短臂的缺失 (5p-)。
■ 主要症状:哭声轻,音调高,很像猫叫;两眼距离宽,耳位低下,智力迟钝,生活力差,多在生命早期死亡。
5.2.3 染色体重复
-
染色体重复 (
duplication) 是指染色体的部分区段出现多个拷贝。对个体的影响相对缺失较为缓和,但大段的重复也会影响生活力。起源于同源染色体之间的断裂和错误重接,或染色单体之间的不等交换等。 -
重复案例:
46, XY, dup(4)(q13q31)
46 条染色体,性染色体组成为 XY,4 号染色体的 q13 至 q31 发生重复。 -
顺接重复 (
tandem duplication) : 染色体某区段按照染色体自身的正常直线顺序重复。如:123456 → 12343456 -
反接重复 (
reverse duplication): 染色体某区段按照染色体自身直线顺序的反向顺序重复。如:123456 → 12344356 -
重复的细胞学效应
■ 同源染色体配对时形成弧状结构/拱环。
■ 弧状结构的内部是重复的染色体部分。
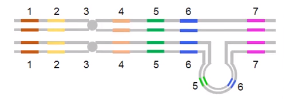
-
案例:果蝇的棒眼基因 Bar

有一种突变果蝇表现出复眼中的小眼减少,所以复眼呈棒状,而不是正常的卵圆形,决定该表型的基因定义为 Bar。对比棒眼果蝇和野生型果蝇的染色体,发现棒眼果蝇 X 染色体上至少有 4 个明显横纹发生了重复;纯合棒眼雌蝇产生的子裔中约有 1/1600 的几率复眼极度细小,叫做重棒眼,检查这种个体的唾腺染色体发现,有 4 个明显横纹的区域被重复了 2 次,说明这个区域的重复是有累加作用的,重复次数增加越多,复眼中的小眼数目就减少越多;可见,棒眼基因的重复带来了显性突变,而重复拷贝数与表型之间是存在关系的。
■ 重复染色体区段内的基因也发生了重复,影响基因表达产物的剂量,所以染色体的重复也会带来突变的表型。
■ 纯合的棒眼雌蝇所产生的子裔中出现了重棒眼的个体的机制与不等交换有关。 -
不等交换 (
unequal crossing over) 即同源染色体在联会时发生错误,在不完全对等的位置上发生交换。不等交换也是造成染色体区段的重复的原因之一。
如下图,纯合棒眼雌蝇产生配子时,同源染色体发生联会,由于 bar 基因所在的 16A 横纹区发生重复,因此可能会出现错误的同源配对并发生交换,交换的结果是,一个配子中 16A 的拷贝数回复正常,另一个配子 16A 有 3 个拷贝,额外重复了 2 次。

-
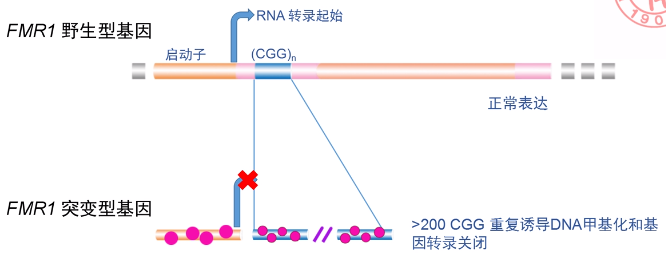
案例:脆性 X 染色体综合征
△ fragile X syndrome 是由于 Xq27.3 染色体重复造成的。
△ 患者智力低下、长脸、大头、大耳、前额突出,下颌大而前突,嘴大唇厚发音障碍。
△ 重复染色体的部分呈收缩或缝隙样,很容易被打断,被称为脆性染色体。脆性 X 染色体既是基因突变,也是一种特殊的染色体畸变。

△ Xq27.3 处有一个编码基因 FMR1,患者的 FMR1 内部 CGG 三核苷酸拷贝数异常增加,正常个体 16-54 个拷贝,患者体内增加到 200 以上甚至上千,CGG 拷贝数异常增加诱导了异常的 DNA 甲基化修饰,进一步使得染色体的转录活性被抑制,从而关闭了 FMR1 的表达。
-
DNA 序列中由于寡核苷酸拷贝数目的变化,引起生物表型改变的突变,称为动态突变 (
dynamic mutation)。动态突变造成的表型变异的程度通常和重复拷贝数的多少有关。又如,亨廷顿舞蹈症。 -
重复与新基因的产生
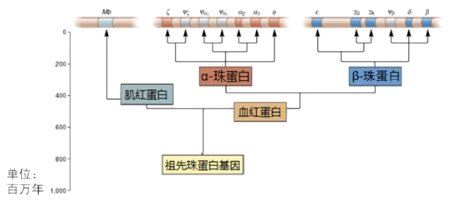
前面的案例重复带来的表型都是不利于生物个体存活的,但从基因组演化的角度,染色体重复起始可以带来基因的重复,从而带来额外的遗传信息,因为是额外的,选择压力小,可以为后来执行新的功能而做出准备。例如,珠蛋白基因的演化,人类基因组中珠蛋白编码基因的拷贝数较多,但在低等生物中,珠蛋白编码基因只有一个,可能是染色体重复促成了血红蛋白和肌红蛋白编码基因的分化,又促成了α-珠蛋白和β-珠蛋白基因的分化。因此,染色体重复带来基因重复,是基因组演化的重要原始材料之一。
5.3 染色体倒位
5.3.1 倒位
-
染色体倒位 (
inversion) 是指染色体一个区段发生了 180 度的颠倒。 -
多数情况下倒位携带者的表型相对正常。
-
倒位起源于单条染色体发生两次断裂后,断裂点之间的片段旋转 180 度后重接。
-
倒位案例:
46,XX,inv(4)(q13q24)
46 条染色体,性染色体组成为 XX,4 号染色体的 q13 至 q24 发生倒位。 -
臂内倒位和臂间倒位
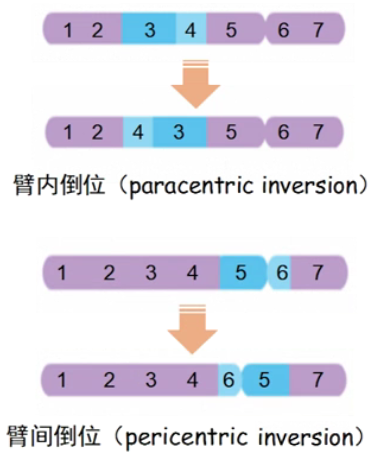
-
倒位与演化
倒位现象存在于很多有亲缘关系的物种中,提示了倒位这种染色体结构变化也与基因组演化之间具有重要的联系。
例如,人和黑猩猩的基因组非常接近,人与黑猩猩的 4 号染色体有一个臂间倒位的差异。

5.3.2 倒位的细胞学和遗传学效应
- 倒位的细胞学效应
倒位杂合子在减数分裂中,为了使发生倒位的染色体与正常染色体能够配对交换,形成独特的倒位环(inversion loop)结构。
但并不是所有的倒位杂合体都会形成倒位环,如果倒位区域较小,只要倒位区域不和标准序列的同源部分配对就可以了。


- 倒位的遗传学效应
倒位的遗传学效应与染色体交换密切相关。
■ 臂内倒位杂合子形成配子,重组型配子中存在桥和断片,可造成配子死亡。
△ 臂内倒位杂合子在联会时,倒位区以倒位环的方式相互配对,如果在倒位环内发生了一次染色体的交换,那么在减数分裂后期时,这两条发生交换的染色单体就会变成另外两种形式,一个含有两个着丝粒,这条有两个着丝粒的染色单体称为称为从一极跨向另一极的双着丝粒桥,另外一个没有任何着丝粒的称为称为断片,这种特殊的染色体组成就叫做"桥和断片”。没有着丝粒的断片因为无法被纺锤体牵引,往往留在细胞质中,不能进入子细胞的核内;而有着两个着丝粒的桥则会被随机的拉断,然后虽然它能进入子细胞的核内,但因为有大程度的重复和缺失,会使配子死亡。
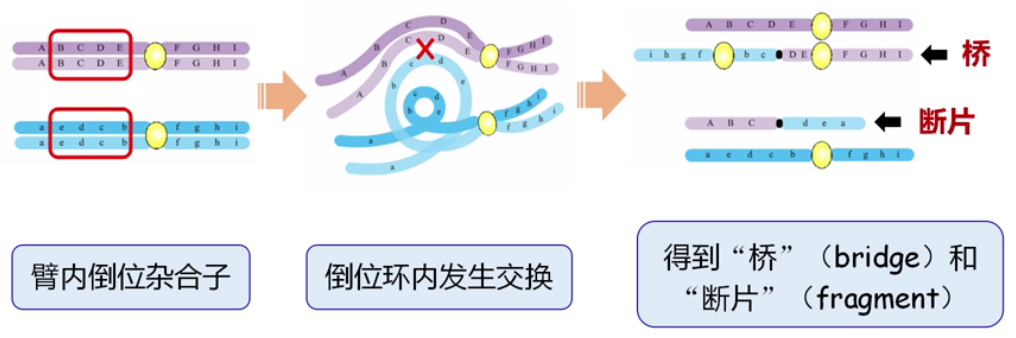
■ 臂间倒位杂合子如果在倒位环内发生交换(交换区域包含着丝粒),不会产生桥和断片,但会产生两种重组的新配子,重组型配子携带重复与缺失,不能存活。

(1) “抑制重组”假象:倒位杂合子在形成配子时,一旦在环内发生交换,就无法得到正常配子,因此,倒位可以抑制或者大大降低倒位环内基因的重组,所以抑制重组是倒位的一种特殊的遗传学效应,但这只是一种假象,事实上重组并非没有发生,只是发生重组之后得到的配子没有办法存活。
(2)倒位发生的具体位置可能会影响到该处的蛋白质编码基因或其他功能性 DNA 的作用,从而造成突变表型,这称之为倒位的位置效应。
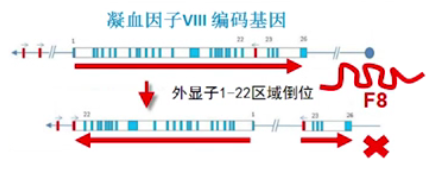
■ 案例 1:倒位与血友病
北美人群中,近 45%的重症甲型血友病家族的发病机制是凝血因子Ⅷ的编码基因外显子 1-22 区段发生倒位,倒位完全破坏了原有的蛋白质编码序列,导致没有凝血因子的正常表达。
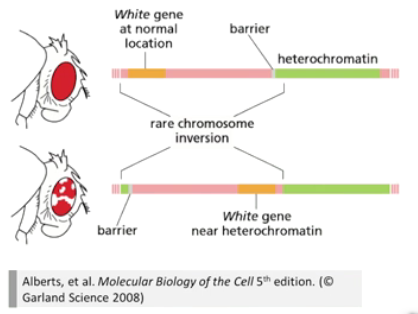
■ 案例 2:倒位与果蝇白眼突变
白斑眼的果蝇突变品系携带 X 染色体倒位。在正常 X 染色体上 w 基因远离异染色质区,可以正常表达,因此是红眼。但在突变果蝇中,由于发生倒位,白眼基因被转移到了异染色质区域旁边,这个区域基因的表达非常低,导致基因沉默,出现了白眼症状。
5.3.3 平衡致死系
-
平衡致死系是倒位的一种重要遗传学应用,是为了解决保留隐性致死基因突变的问题而设计的。
-
果蝇的一般品系都以纯合子的形式进行保存的,但是致死基因不能以纯合子的状态保存,只能以杂合的状态进行保存。
-
隐性致死等位基因的保存
例如,翘翅基因 (Cy): 显性性状基因,翻翅,也是纯合致死基因。
为了保存这一品系,只能把 Cy + 杂合子果蝇自交,得到的子一代是 2/3 Cy + 杂合子,和 1/3 ++ 野生型果蝇(CyCy 纯合子死亡,因此不计入后代统计)。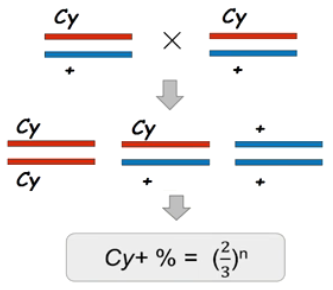
由于每代都会产生 1/3 的野生型纯合果蝇,n 代后,这个杂合子的比例就会变成 ( 2 3 \frac {2}{3} 32)n,如果 n 趋近于∞,( 2 3 \frac {2}{3} 32)n 就趋近于 0,那么 Cy 基因就遗失了。因此,每一代都要观察后代,把野生型个体淘汰掉,但这样工作量太大,一个巧妙的方法是,用致死基因“平衡”另一致死基因。 -
平衡致死系
■ 平衡致死系 (Balanced lethalsystem): 用非等位基因的双杂合子保存隐性致死基因的品系。两个非等位基因,任何一个出现纯合,都是致死的,只有两个基因都为杂合状态的个体才能够保留下来。同时,利用倒位片段抑制交换重组,保证杂合状态在世代传递中不发生分离。
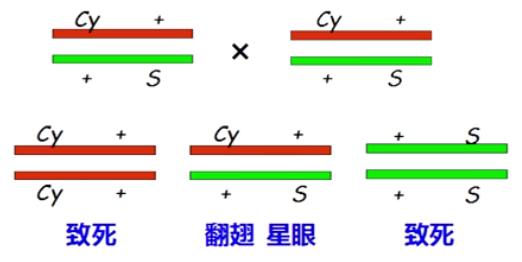
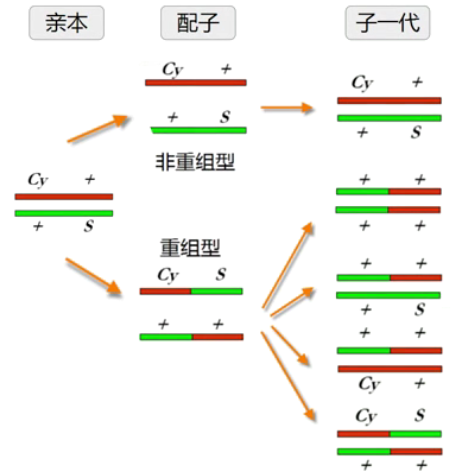
■ 先把 S+杂合子与 cy+杂合子杂交,可以得到反式杂合子(S+/+Cy),然后把这个反式杂合子继续自交,后代仍然保持 S+/+Cy,因为后代中的 SS 和 GyCy 纯合子都死亡了,只有杂合子保留下来,且不会发生分离。这种永远以杂合状态保留下来,不发生分离的品系就是永久杂种/平衡致死系。
■ 但还有一个要求:不能发生重组交换。一旦发生重组,就会丢失 Cy 或 S,Cy 和 S 就不能继续保存下去。利用倒位片段抑制交换重组。例如,果蝇第 2 染色体上倒位品系 Cy 在ⅡL(第 2 染色体左臂)有个倒位,在ⅡR(第 2 染色体右臂)也有个倒位,几乎把整个第 2 染色体的交换全部抑制。而 Cy 的纯合是致死的,因此第 2 染色体上任何致死基因都可用 Cy 来平衡。
小结:
- 倒位没有影响遗传物质的量,但改变了遗传物质的排列顺序。
- 倒位杂合子可以形成特殊的倒位环结构,而且具有抑制重组的遗传学效应。
■ 思考:如果倒位环内发生的是双交换,配子的遗传组成将会是什么?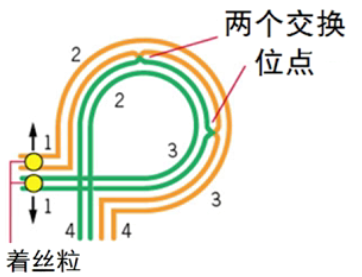
- 平衡致死系可以用来长期保存隐性致死突变。
5.4 染色体易位
5.4.1 易位
- 染色体易位 (translocation) 是指染色体片段的位置转移。多数情况下易位携带者的表型相对正常。
- 易位通常起源于两条非同源染色体的断裂和错误连接。
- 相互易位 (reciprocal transl-ocation) 指的是两个非同源染色体互相交换染色体片段。相互易位只改变位置关系而不造成遗传物质丢失的现象。
案例:46,XY,t(4; 20)(q25; q12)
46 条染色体,性染色体组成为 XY,4 号染色体的 q25 至未端,与 20 号染色体的 q12 至末端之间发生相互易位

- 罗伯逊易位(Robertsonian translocation),又称着丝粒融合 (centric fusion),是发生在两对近端着丝粒染色体之间的一种易位形式。可以形成一条由两条染色体长臂构成的衍生染色体,由两个短臂构成的染色体容易在后续分裂中发生丢失,会导致染色体数目的变异。罗伯逊易位携带者表型一般也正常,因为丢失的染色体片段小,丢失的遗传物质不多,但在携带者生殖细胞中,由于染色体数目变异,配子在形成时发生异常。
- 易位与演化
人类染色体组有 23 对 46 条染色体,而黑猩猩染色体组是 24 对 48 条,对比后发现,人类的 2 号染色体长臂和短臂分别和黑猩猩的两条染色体之间存在明显的同源关系,因此,人类的 2 号染色体可能是由祖先的两条独立的染色体发生罗伯逊易位产生的。在包括黑猩猩在内的现代类人猿中,都能着到未发生易位融合的两条独立染色体,可见,易位也可以促进基因组的演化。

5.4.2 易位的细胞学与遗传学效应
- 相互易位杂合子在形成配子时,共四条染色体彼此配对在一起,形成特殊的“十”字结构。
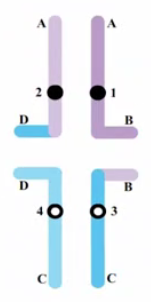
- 在生殖细胞形成过程中,易位会对配子的存活率和非等位基因的连锁关系造成影响。由于相互易位杂合子的同源染色体联会涉及到了 4 条染色体,并形成了十字形,易位出现了两种特殊的遗传学效应:半不育现象和拟连锁。
(1)半不育现象 (
semisterility)- 减数分裂第一次分裂后期,同源染色体分离,由于联会的相互易位染色体有 4 条,在分离时可能会采用不同的分离方式:
① 邻近分离:得到的两种类型配子含大量重复和缺失,是不平衡的,常有致死效应,不育。
② 交互分离:得到两种新类型配子,其中一个配子内含有两条正常染色体,另一个配子含有两条易位染色体,虽然携带了易位染色体,但是染色体是平衡的,没有重复或缺失,可育。 - 因此,有且仅有交互分离方式下,可以得到可存活的配子,这就是半不育现象。
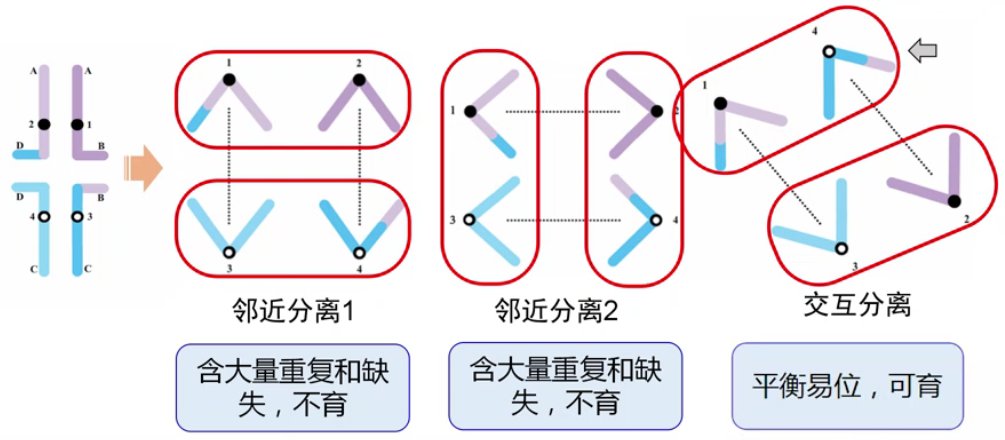
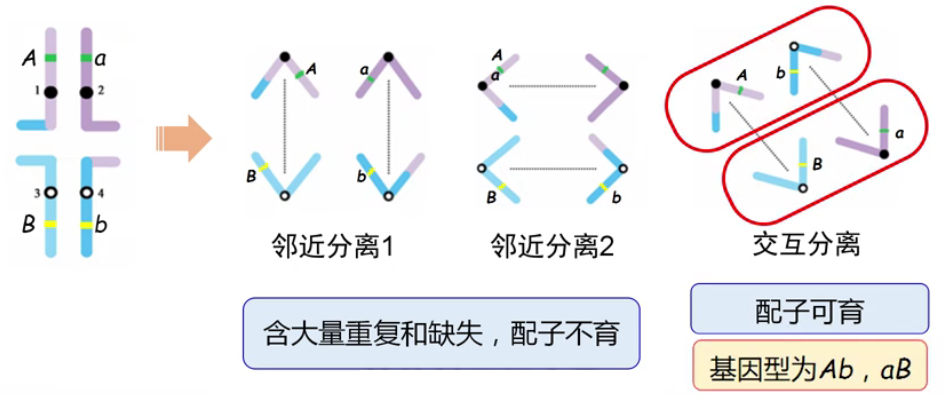
(2)拟连锁 (
pseudolinkage)-
这种效应的产生与配子半不育之间也有密切联系
-
假设发生相互易位的 4 条染色体上分别携带了不同的等位基因 A、a、B、b,A 和 B 是不同染色体上的基因,他们之间一定不连锁,是自由组合的关系。由于只有发生交互分离的配子可存活,因此,AaBb 的亲本只能得到两种类型配子:Ab、aB。这种结果从表面现象看,Ab 之间、aB 之间是连锁的,这种连锁的假象称为拟连锁,它实际上是由于部分配子类型不能存活所造成的。
-
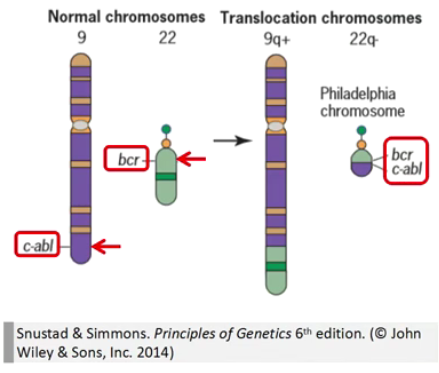
体细胞携带的染色体易位有时也会带来严重的表型效果,往往和易位的断裂点位置有关,与倒位类似。
案例:易位与肿瘤
慢性髓系白血病 (CML) 中 chr 9(c-ABL) 与 chr 22(BCR) 发生相互易位,产生组成型激活的 BCR-ABL 融合蛋白,可以脱离调控直接刺激细胞的快速分裂与增殖。
利用 BCR-ABL 融合蛋白的抑制剂 Gleevec(Imatinib),结合 BCR-ABL 的 ATP 结合位点,抑制 BCR-ABL 重组蛋白的活性,从而治疗携带这种易位染色体的晚期 CML 患者。临床试验显示,Gleevec 的肿瘤抑制效果非常显著,药物治疗后 5 年生存率可达 80-90%。
5.4.3 其他染色体结构变异
-
环状染色体 (
ring chromosome) 指的是闭合成环的染色体。起源于单条染色体的长、短臂断裂后,含有着丝粒的中间片段的两端的错误重接。
环状染色体案例:46, XY, r(2)(p21q31)
46 条染色体,性染色体组成为 XY,2 号染色体的 p21 至 q31 形成闭合环。

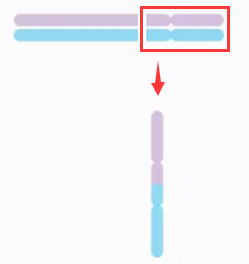
-
双着丝粒染色体 (
dicentirc chromosome) 指的是具有两个着丝粒的变异染色体。起源于两条染色体的断裂后,两个具有着丝粒的片段重接。
双着丝粒染色体案例:46, XX, dic(5;9)(q31;q21)
46 条染色体,性染色体组成为 XX,5 号染色体的 q31 至未端和 9 号染色体的 q21 至未端形成双着丝粒染色体
-
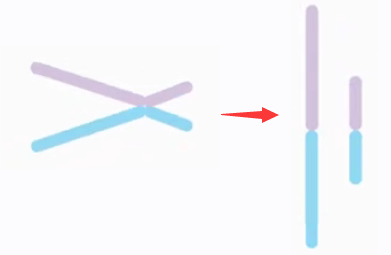
等臂染色体 (
isochromosome) 指的是一条染色体的两条臂在形态和遗传结构上完全相同。起源于细胞分裂中着丝粒异常的横裂。
等臂染色体案例:46,XX,i(12q)
46 条染色体,性染色体组成为 XX,12 号染色体的长臂形成等臂染色体
-
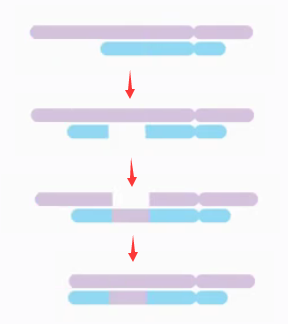
插入 (
insertion) 指的是一条染色体的片段插入到另一条染色体中的现象。起源于两条染色体的三次断裂和错误重接。
插入案例:46, XY, ins(18;5)(q21;q31q35)
46 条染色体,性染色体组成为 XY,5 号染色体的 q31 至 q35 区段插入到 18 号染色体的 q21 位置
-
染色体结构变异的分子机制
Liu, P et al.Curr Opin Gen-et Dev. 2012
① 非同源末端连接 (nonhomologous end-joining, NHEJ) 是真核生物细胞在不依赖 DNA 同源性的情况下,而为了避免 DNA 或染色体断裂的滞留,避免因此造成的 DNA 降解或对生命力的影响,强行将两个 DNA 断端彼此连接在一起的一种特殊的 DNA 双链断裂修复机制。NHEJ 是一种基因组常采用的修复 DNA 双链断裂的方式,修复中的错误往往会导致畸变的发生。
②非等位同源重组 (non-allelichomologous recombination, NAHR),指的是两条同源的但在基因组不同位置重复出现的高度相似的 DNA 序列之间的配对和交换(不等交换)。基因组中的大量重复序列之间可以通过非等位同源重组,产生染色体的结构变异。
③ DNA 复制机制 (replicative mechanism) 包括基于复制叉停滞与模板交换等具体机理。
5.5 染色体的数目变异——单倍体与多倍体
5.5.1 染色体数目变异
- 任一生物的染色体数目都是恒定的,染色体数目的变异和结构变异一样,也是染色体畸变的重要组成部分。
- 一个配子内的全部染色体构成一套完整的染色体组。体细胞内染色体数目是染色体组整数倍的生物个体被称为整倍体 (
euploid)。 - 含有两套完整的染色体组的细胞核型是二倍体 (
Diploid)。如 Diploid(2n=6),表示单套染色体组是三条染色体的个体。 - 含有单套染色体组的细胞核型称为单倍体 (
Haploid)。如 Haploid(n=3)。单倍体就是一种染色体的数目变异。 - 含有超过两套染色体组的细胞核型称为多倍体 (
Polyploid)。如 Polyploid(3n=9)。 - 如果体细胞的染色体组中的一条或几条染色体发生改变,这种非整数倍的改变形成非整倍体 (
aneuploid)。如 aneuploid(2n-1=5),二倍体缺一条染色体是单体 (2n-1),二倍体多一条染色体是三体 (2n+1)。
5.5.2 单倍体
- 单倍体生物的细胞内只存在单套染色体。
- 一些低等植物的配子体和一些膜翅目昆虫的雄体是天然的单倍体。如雄蜂是由未受精的卵细胞经孤雌生殖发育而来的。
- 单倍体生物的遗传学效应
■ 正常单倍体形成的配子仍为单倍体。
■ 单倍体变异细胞在进行减数分裂时,没有可以配对的同源染色体。所有染色体都是随机地被纺锤体牵引到细胞的一极
■ 单倍体变异细胞产生可育配子的概率很低,所有的染色体都分到一极的概率是 ( 1 2 \frac {1}{2} 21)n-1(n 是单倍体的染色体数)。 - 单倍体与作物育种
■ 花药离体培养法是一种常见的利用单倍体细胞/组织进行育种的方法。将花药进行体外的培养,诱导成单倍体植株,再通过人工诱导使得染色体数的目加倍,恢复到正常染色体数目,因此,花药离体培养得到的二倍体植株的每对染色体上成对的基因都是纯合的,自交产生的后代不会发生性状分离,有利于隐性纯合突变植株的建立,比常规的杂交育种育种年限短。
■ 但花药离体培养产生单倍体的技术不是对所有生物或所有基因型都适用,且这种方法还利用了植物细胞的全能性和植物激素的染色体加倍作用这两个重要性质,因此这种方法不适用于动物。
5.5.3 多倍体
- 多倍化和杂交是高等植物基因组进化和新物种形成的主要动力之一。
- 约 70%的被子植物的种类具有多倍体 (Masterson,1994)。多倍体植物在栽培作物、果树、蔬菜中尤为常见。因为,在一般情况下,多倍体的物种在营养、产量、品性或者观赏价值和食用便捷程度上相对二倍体是有很大优势的,比如常见的棉花、小麦、草莓、香蕉、咖啡、玫瑰、菊花等。
- 动物中罕见多倍体,多倍体动物包括美洲角蛙(4 倍体)、银鲫(3 倍体)、牡蛎(3 倍体)等。
- 多倍体可以根据起源的机制不同分为同源多倍体和异源多倍体。
同源多倍体
- 同源多倍体 (
autopolyploid) 产生于一个物种的染色体组加倍,加倍前后染色体种类相同,数目不同。
■ 例如,原物种 2n=10,染色体复制但细胞质没有发生分割,未减数的配子结合之后就可以发育成同源多倍体 4n=20(四倍体)。
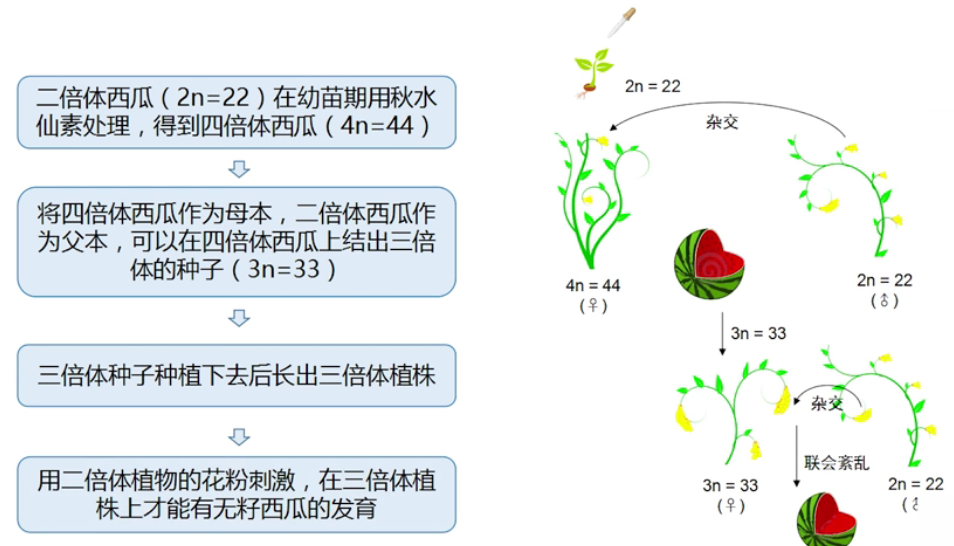
■ 在同源多倍体中,所有的基因和原来相同,只是拷贝数不同。 - 案例:三倍体无籽西瓜
- 同源多倍体的遗传学效应
■ 同源多倍体细胞在联会时,会有三条或以上的染色体参加联会,形成“多价体”(mult-ivalent)。如∶三价体 (Ⅲ)、四价体 (Ⅳ)。
■ 同源三倍体的联会和分离
联会时会有两种可能:
① 形成一个二价体和一个单价体,二价体同源染色体分离,分别进入细胞的两极,而单价体将随机进入细胞的一极。如果每个单价体都进入同一极,就可以得到一个单倍体配子和一个二倍体配子,但这种可能性很低,多数情况下,得到的是 n+X 条染色体,即非整倍体。
② 形成三价体,然后同源染色体随机分离,2/1 分离或 3/0 分离,如果每组同源染色体的分离方向都一致,就能得到单倍体、二倍体和三倍体的配子细胞,如果分离方向是随机的,就会得到非整倍体(可能性更大)。
因此,无论是哪种联会方式,三倍体形成的配子染色体组的组成往往不平衡,可育性很低。

■ 同源四倍体的联会和分离
同源四倍体采用的分离方式有三种:
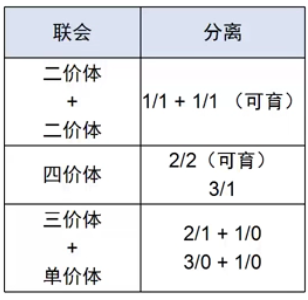
因此,同源四倍体减数分裂形成的配子部分可育,但多数是非整倍体,不可育。
异源多倍体
-
不同物种杂交,杂种再经过染色体加倍可形成异源多倍体 (
allopolyploid)。异源多倍体的染色体数目不同,且种类不同。 -
例如,2n=8 的物种和 2n=6 的物种杂交,得到 2n=7 的子代,由于所有染色体均不同源,不能产生正常的配子,所以需要经过植物激素的刺激使染色体组加倍,得到 4n=14 的异源多倍体。这个多倍体在减数分裂时,联会和分离的情况更像一个二倍体,同源染色体形成的都是二价体。

-
异源多倍体是生物演化、新物种形成的重要因素之一。
-
许多农作物是人工改良的异源多倍体。
-
利用染色体组分析和人工杂交实验可以揭示异源多倍体的起源。
-
案例:普通小麦
将所有要分析的不同小麦之间进行杂交,观察杂种在减数分裂过程中染色体的配对情况,结果如下表:

(1)第 1 个亲本组合的配对情况是 7 个二价体和 7 个单价体,用 AA 表示 7 个二价体中对应的染色体组,B 表示 7 个单价体所对应的染色体组,那么一粒小麦的染色体组组成可能就是 AA,二粒小麦的染色体组组成可能就是 AABB;
(2)第 2 个亲本组合配对情况说明,一粒麦草和滔氏麦草的染色体组没有同源关系,滔氏麦草的染色体组是非 A 的;
(3)第 3 个亲本组合配对情况说明,二粒麦草和滔氏麦草的染色体组之间也没有同源关系,因此滔氏麦草的染色体组用 D 表示;
(4)第 4 个亲本组合配对情况说明,普通小麦中含有 7 条和一粒小麦同源的染色体组 A;
(5)第 4 个亲本组合配对情况说明,普通小麦中含有 14 条和二粒小麦同源的染色体组 AB;
(6)第 4 个亲本组合配对情况说明,普通小麦中含有 7 条和滔氏麦草同源的染色体组 D;
综上,综普通小麦的单套染色体组的组成就是 ABD,二倍体的组成就是 AABBDD。
5.6 非整倍体
5.6.1 非整倍体概念与分类
-
非整倍体 (
aneuploid) 是生物个体染色体组的一条或几条染色体数目的非整倍性改变形式。 -
超二倍体 (
hyperdiploid) 比二倍体染色体组多一条或几条染色体。 -
亚二倍体 (
hypodiploid) 在二倍体染色体组少一条或几条染色体。 -
单体:属于亚二倍体
■ 正常二倍体缺少一条染色体称为单体 (monosomy) (2n-1)。
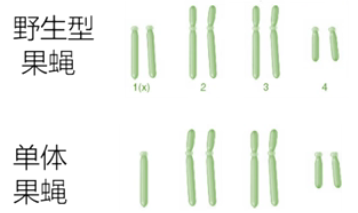
■ 注意与单倍体区分,单倍体是 n。
■ 大多数动植物的单体都不能存活,缺少一条染色体常常比缺少一套染色体对个体生存的威胁更大,说明不同染色体所携带的遗传物质相互平衡的重要性。
■ 自然界中也有少数天然的单体。如,黑腹果蝇和线虫等生物中,缺少一条 X 染色体的单体是正常的雄性;小麦的单体植株表现型很不正常,但能存活。 -
缺体:属于亚二倍体
■ 缺失了一对 同源染色体的个体称为缺体 (nullisomy) (2n-2)。

■ 缺体对生物个体的性状表现的影响更大,缺体的生活力更差。如,普通烟草的缺体在幼胚阶段就会死亡。
■ 但也有可存活的天然缺体,如小麦的 21 种缺体都是可存活的。
■ 注意:如果某突变体丢失了 2 条非同源的染色体,叫做双单体,而非缺体,表示为2n-1-1。 -
三体:属于超二倍体
■ 在二倍体中增加了一条染色体形成的突变体称为三体 (trisomy) (2n+1)。

■ 三体对个体的影响比单体小,但是增加的染色体也会使个体出现显著的形态和生理变化,并导致配子不育。
■ 自然界中有天然三体,如曼陀罗有 12 种三体类型,恰巧和曼陀罗的单倍体染色体数目 n=12 是相同的。
5.6.2 非整倍体与人类疾病
21 三体综合征
- 又称唐氏综合征,新生儿中发病率约为 1/650。
- 患儿具有特殊的面貌,头颅前后径短,枕骨扁平,眼小,两眼外侧高而内侧低,鼻梁扁平且宽,口半张,舌常伸出口外,舌有龟裂。掌纹和指纹特殊,常为通贯手。发育迟缓,智力低下。平均寿命很短,大约到 10 岁时已有 1/3 患者死亡。
- 1959 年,Lejeune 等人揭示唐氏综合征的发病机制是人类的 21 号染色体多出了一条。

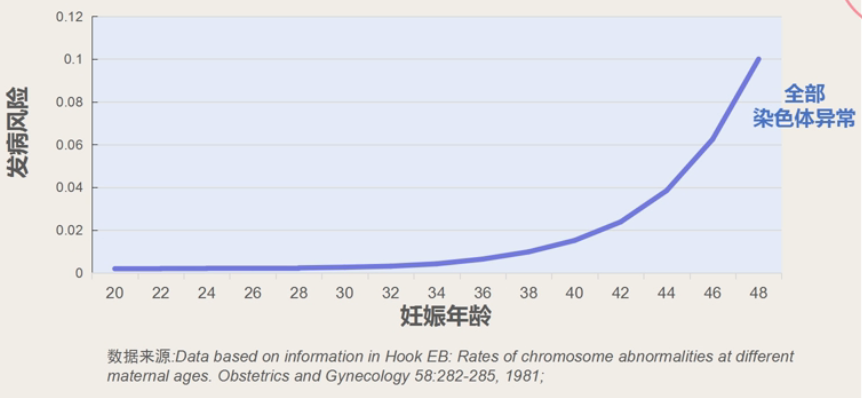
- 高龄孕妇生育唐氏综合征患儿比例偏高
数据显示,年龄在 25 岁以下的孕妇生育唐氏患儿的概率很低,约 1/1500,35 岁以上的孕妇生育患儿的概率可提高至约 1/250,且年龄越大概率越高。
携带各种染色体变异的全部患儿总数也随着孕妇年龄的上升而逐渐升高。35 岁以生育患病孩子的概率显著高于 35 岁之前。临床统计分析还发现,如果将≥35 岁的孕妇年龄单独作为筛查指标,检出率为 32%。
- 唐氏综合征目前尚无有效治疗方法,因此,开展唐氏综合征的产前筛查和诊断,通过终止妊娠来防止患儿出生是目前唯一有效的干预措施。
- 产前标志物筛查∶孕妇年龄、孕妇血清标志物、超声测量等。根据各种已证明的与唐氏综合征相关的筛查标志物来估计孕妇怀有唐氏综合征患儿的风险。
- 产前核型诊断:核型分析。如果产前标志物指示风险较高,就需要进一步确诊,做羊水穿刺,获得胎儿的脱落细胞,再利用核型分析确定胎儿是否携带了 3 条 21 号染色体。风险:羊水穿刺容易引发胎儿流产。
- 新的诊断方法:胎儿游离 DNA (cfDNA) 检测。只需要孕妇的血浆,分析其中的 cfDNA 即可。相比传统的血清学方法,准确性和灵敏性都有所提高;相比核型分析,无流产风险。
18 三体综合征
- 18 三体综合征 / Edward 综合征

- 发病率约 1/8000。临床表征:身体、智力发育迟滞,头长,多种器官先天畸形,口鼻小等,早天。
13 三体综合征
- 13 三体综合征 / Patau 综合征

- 发病率约为 1/20000。临床表征︰患者严重智力迟钝、小头畸形,先天性心脏病等,早夭(常在出生 3 个月内死亡)。
- 在出生缺陷的患儿中,有且仅有以上三种常见的常染色体非整倍体,其他常染色体的三体仅见于先天流产的胚胎中,说明相对于其他常染色体,13、18,尤其是发病率最高的 21 号染色体其实反而是发育过程中,相对不重要的染色体。
- 在出生缺陷的患儿中,没有常染色体单体,说明缺少一条染色体比增加一条染色体对个体生活力的影响更大。
- 相对于常染色体,性染色体的重复或缺失对个体的影响力较小,通常只影响生殖能力,而与存活率之间没有联系。说明,性染色体在基因组中的作用相对较小。
非整倍体的形成机制
● 以 21 三体为例,造成 21 三体综合征的机制包括:
-
减数分裂中染色体不分离
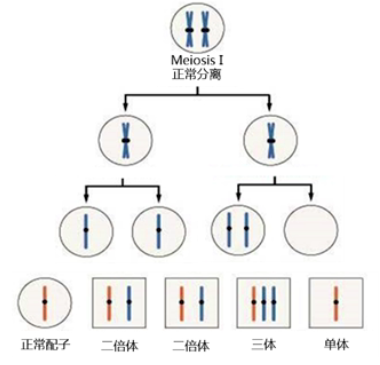
■ 染色体不分离 (nondisjunction) 指的是同源染色体或者是姐妹染色单体在分裂后期不发生分离,同时移动到细胞同一极的现象。
■ 减数分裂不分离可以发生在减数第一次分裂、减数第二次分裂后期。
(1)当不分离发生在减数第一次分裂后期时,配对的同源染色体不分离,而是被分配到同一个次级母细胞中,在成熟配子中则将有 2 个配子含有 n+1 条染色体,另外 2 个含有 n-1 条染色体,它们和正常配子结合后得到 2 个三体和 2 个单体。
(2)当不分离发生在减数第二次分裂后期时,同源染色体正常分配到次级母细胞中,随后在每一个次级母细胞中,出现姐妹染色单体不分离,被分到同一个配子中,因此,在成熟配子中,有 2 个配子是正常野生型配子,另外 1 个含有 n+1 条染色体的三体和含有 n-1 条染色体的单体,它们和正常配子结合后得到 2 个正常二倍体、1 个三体和 1 个单体。
■ 如果在卵细胞发育过程中,出现染色体不分离,异常的卵细胞与精子结合后就可能得到三体。反之,如果精子异常,也有可能得到三体。但临床调查发现,造成 21 三体综合征的主要原因是卵细胞的异常,少数是精子异常。这和女性生殖细胞的特殊发育过程是密切相关的,减数分裂实际上是在女性还是胎儿的时期就已经开始,直到受精卵受精时才真正完成,因此在受精完成之前的几十年中,减数分裂细胞一直停滞在减数第一次分裂的前期,容易积累错误,显然,停滞时间越长,错误积累的可能性越大。 -
有丝分裂中染色体不分离
■ 染色体不分开现象也可出现在合子形成以后的有丝分裂过程中。
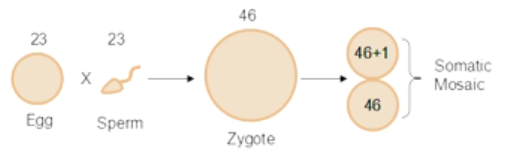
■ 如果出现在卵裂期或以后,就有可能形成由不同基因型细胞所组成的个体,也就是嵌合体。但是 21 三体综合征嵌合体临床上不常见,常见的是性染色体的嵌合体,在这些个体身上可以同时具有男性和女性的组织。如,嵌合体组成是 XO/XYY,就是 XY 合子中 Y 染色体不分离产生的。 -
罗伯逊易位
■ 表型正常的罗伯逊易位携带者也有可能生育 21 三体患儿。
■ 如,一个表型正常的男性,携带发生罗伯逊易位的染色体,14 号短臂与 21 号染色体长臂融合,染色体数是 45,但染色体总量是平衡的,因为其中一条融合的 14 号染色体上包含了 14 号和 21 号染色体各一条。
■ 但当该男性的生殖细胞发生减数分裂时,融合的染色体 T(14,21) 就可以和正常的 14、21 号染色体形成三价体,三价体的随机分离可能出现 3 种结果,得到 6 种配子,这 6 种配子和一个正常卵子结合后就会得到不同的子代,分别是 14 单体(死亡)、14 三体(死亡)、21 三体(患者)、21 单体(死亡)、野生型二倍体以及易位平衡携带者(与父亲的情况相同),如下图。

- 细胞中染色体的总体构成称为核型 (
-
相关阅读:
50 二叉树中的最大路径和
是否在业务中使用大语言模型?
Java 并发编程解析 | 如何正确理解Java领域中的并发锁,我们应该具体掌握到什么程度?
MySQL技术内幕-InnoDB存储引擎
软件加密系统Themida应用程序保护指南(四):虚拟机的选择
057_末晨曦Vue技术_处理边界情况之强制更新和创建低开销的静态组件
ubuntu 22 安装 python3.11.7
Mybatis-Plus使用Wrapper自定义SQL
Python 轻松生成PDF文档
Wireshark TS | MQ 传输缓慢问题
- 原文地址:https://blog.csdn.net/zea408497299/article/details/126072804